


| name | Amanita virosa |
| name status | nomen acceptum |
| author | (Fr.) Bertillon in Dechambre |
| english name | "European Destroying Angel" |
| images |
 1. Amanita virosa, Brekke sluser, Halden, Ostfold, Norway.  2. Amanita virosa, Degerberget, Hörnefors, Västerbotten, Sweden.   3. Amanita virosa, Degerberget, Hörnefors, Västerbotten, Sweden. |
| intro |
The following is largely based on the descriptions by Bresinsky and Besl (1990) and Neville and Poumarat (2004). |
| cap |
The cap of Amanita virosa is 29 - 123 mm wide, white, sometimes pale cream-colored, sometimes with yellowish or pale orangish tan tints in the center with age, hemispheric when young, soon conico-campanulate, with a broad umbo, smooth, viscid when moist, shiny when dry, not symmetric (with an irregular shape, not circular, often lobed), with a nonstriate and nonappendiculate margin. The volva is usually absent but rarely occurs as a few white membranous patches. The flesh is white, thick in the center of the cap, thin towards the margin. |
| gills |
The gills are quite close, pure white to cream, with a flocculose edge. The short gills are truncate. |
| stem |
The stem is 50 - 165 × 7 - 15 (-20) mm, cylindrical, white, solid to pithy-hollow, scaly below the ring (often illustrated with recurved pointed scales), arranged in concentric rings and somewhat overlapping each other; in some cases the scales are robust. The bulb is 16 - 48 mm wide. The ring is white to yellowish, skirt-like, membranous, fragile, collapsing rapidly on the stem, and in the upper quarter of the stem. The volva is membranous, white, sometimes taking on a pinkish tint at maturity, arising from the upper surface of the bulb, limbate, and usually collapsing against the stem base. The flesh is pure white and unchanging. |
| odor/taste | The odor is strongly of a old rose or honey, in age or drying. This species is deadly POISONOUS. |
| spores |
Bresinsky and Besl (1990) measured spores as follows: 8.2 - 11.3 × 6.7 - 9.7 µm and are subglobose to broadly ellipsoid and amyloid. Clamps are absent at bases of basidia. Neville and Poumarat (2004) measured spores as follows: 8 - 11 × 7.5 - 10 µm and are globose to subglobose to broadly ellipsoid and amyloid. RET measured spores from material collected from France, Norway, and Switzerland as follows: (6.6-) 8.2 - 10.5 (-13.0) × (6.1-) 6.9 - 9.5 (-12.6) µm. |
| discussion |
Amanita virosa is deadly poisonous. This species turns a beautiful and bright yellow on all surfaces when exposed to a 10% KOH solution. Amanita virosa was originally described from Sweden and is known from Europe and eastern Asia. Neville and Poumarat report this species under beech (Fagus sylvatica), chestnut (Castanea satiba), pine (Pinus), spruce (Picea abies), and fir (Abies alba). [Ed. Note: Although the present species has been reported from eastern Asia and has been confirmed genetically there (e.g., in Jilin Province, China), the name has apparently been misapplied to a number of different species. For example, see below.—Zhu L. Yang and RET] Very similar white "destroying angels" that stain yellow with KOH and have dominantly subglobose to broadly ellipsoid spores exist in much of the northern hemisphere: Amanita subjunquillea var. alba Zhu L. Yang and A. exitialis Zhu. L. Yang & T. H. Li (eastern Asia) and A. bisporigera G. F. Atk. and A. suballiacea (eastern North America).—R. E. Tulloss and L. Possiel |
| brief editors | RET |
| name | Amanita virosa | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| author | Fr. ex Bertillon in Dechambre. 1866. Dict. Encycl. Sci. Médic.: 497. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| english name | "European Destroying Angel" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| synonyms |
≡Agaricus (Amanita) virosus Fr. nom. inval. 1838. Epicr.: 3. [Posterior homonym. ICBN §53.1] =Agaricus virosus Vitt. nom. inval. p.p. 1835. Descr. Fungh. Manger.: 135, pl. 17. [Posterior homonym. ICBN §53.1] [Misapplication in part—concept including both A. verna and A. virosa.] non Agaricus virosus Sowerby. 1797-1805. Col. Fig. of English Fungi or Mushro. : pl. 407, 408. [ affin. Stropharia semiglobata (Batsch : Fr.) Quél.] [per Neville and Poumarat (2004: 591)]
For more extended synonymy see the Amanita Nomenclator (t.b.d.). The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MycoBank nos. | 163692 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| neotypes | Fries, E. M. 1861. Sveriges Svampar: pl. 84. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| neotypifications | Neville and Poumarat. 2004. Fungi Europaei 9: 593. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| intro |
Olive text indicates a specimen that has not been
thoroughly examined (for example, for microscopic details) and marks other places in the text
where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of Dr. Z. L. Yang or another researcher is based upon original research by R. E. Tulloss. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
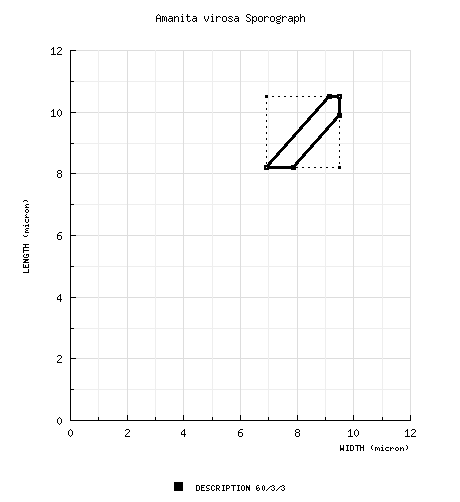
| basidiospores | [60/3/3] (6.6-) 8.2 - 10.5 (-13.0) × (6.1-) 6.9 - 9.5 (-12.6) µm, (L = 8.9 - 9.5 µm; L’ = 9.3 µm; W = 8.4 - 8.6 µm; W’ = 8.6 µm; Q = (1.02-) 1.04 - 1.15 (-1.20); Q = 1.06 - 1.10; Q’ = 1.09), ??, smooth, ??, amyloid, subglobose, occasionally globose, rarely broadly ellipsoid, occasionally pip-shaped, adaxially flattened; apiculus sublateral, cylindric, sometimes proportionately small; contents ??; ?? in deposit. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ecology | Solitary to subgregarious. Norway: In conifer forest. Switzerland: At 400 m elev. With Fagus & Abies. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| material examined |
Zhang et al. (2010) voucher for sequencing: CZECH REPUBLIC: UNKN.—Unkn. loc., s.d. unkn. coll. s.n. (HKAS 55298). Cai et al. (2014): CHINA: JILIN—Changbai Mtn., unkn. loc., s.d. unkn. coll. s.n. (HMJAU 20396). Fusong, unkn. loc., s.d. unkn. coll. s.n. (HMJAU 23303). FINLAND: EASTERN FINLAND—Juva, s.d. unkn. coll. s.n. (HKAS 56694). JAPAN: HOKKAIDO—unkn. loc., s.d. unkn. coll. s.n. (HKAS 71040). RET: FRANCE: ILLE-ET-VILAINE—Brocéliande, x.1998 C. Ricard s.n. [F. Massart 98025] (in herb. Massart; RET 291-3, nrITS & nrLSU seq'd.). NORWAY: OSTFOLD—Halden, Brekke sluser, 14.ix.1975 G. Gulden 302/75 (O 53862). SWEDEN: VÄSTERBOTTEN—Hörnefors, Degerberget, 4.viii.2010 Irene Andersson s.n. [mushroomobserver #49609] (RET 451-6). SWITZERLAND: ZÜRICH—Zürich, 18.viii.1991 C. Lavorato 910818--- (in herb. Lavorato; RET 033-2). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| discussion |
For purposes of comparison, the sporograph (green
figures) for A. bisporigera is presented
here: To date, none of the white "destroying angels" of the eastern United States for which an nrITS sequence has been obtained, have been a good match for the true Eurasian A. virosa. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.